Ecological Wood Adaptation and Horizontal Variations of Vessel Element and Fibre Length of Calligonum mongolicum
Shumin Yang
Shumin Yang*
Research Institute of Wood Industry,Chinese Academy of Forestry,Beijing 100095,China
Abstract
Wood anatomy of Calligonum mongolicum Turcz. was described from an ecological perspective. This species showed similar wood structure to that species published in the same genus [1]. This species has distinct growth ring boundaries, ring-porosity, distinct helical thickenings, simple perforation plate, nonseptate fibre, axial parenchyma, uni- or 2-5-seriate heterogeneous rays, and alternate intervessel pitting. It was observed that there is a slightly increase of fibre length as the diameter from the pith increases. However, the vessel element length remained more or less constant from pith to bark. Furthermore, the relationships between anatomical features and adaptability to desert environments were discussed.
Keywords
Desert shrubs,Ecological wood anatomy,Polygonaceae.
1. Introduction
The larger genus in Polygonaceae is Calligonum,which includes about 100 species of shrubs that grow in central Asia. It is well suited to arid climates with drought resistance,and grows more on clay,sandy and gravel grounds. These plants are often cultivated as ornamentals and a stabilizer of mobile sand dunes.
There is published information on the wood anatomy of all the examined genera in Polygonaceae [1],but the selected species have not yet been described. Additionally,previous studies on wood anatomy of other species have mainly focused on general description [2]. The results of this paper are to provide preliminary information of the wood anatomy of selected species and discuss the relationship between anatomical characteristics of secondary xylem and adaptability to arid climate. Furthermore,the horizontal variations in vessel element length and fibre length are studied. The information obtained through this study will be useful for selecting and introducing suitable species to control desert expansion.
2. Materials and Methods
Five healthy trees were felled and two discs (2-3cm thick) from each tree were taken at the height of 20-30cm above the ground. Some of the discs were immediately fixed in formalin-acetic-alcohol (5:5:90 v/v).
Wood samples were softened in 5% glycerin solution,subsequently sectioned with a sliding microtome moving on transverse,radial and tangential surfaces of the disks. Thin sections were stained with safranine,dehydrated in a graded alcohol series and mounted in Canada balsam for light microscope examination. Small blocks exposing transverse,radial and tangential surfaces were respectively prepared according to Exley et al’s methods [3] for scanning electron microscope (X-650,Hitachi Ltd Tokyo,Japan) observations. Maceration was prepared through soaking in Jeffrey's solution and mounted in glycerin-jelly. Quantitative data were based on 25 measurements of vessel element length and 50 of fibre length. Terminology and methodology followed the IAWA list of microscopic features for hardwood identification [4].
3. Results
3.1 General wood anatomical descriptions
Growth ring boundaries are undulating and distinct,marked by thick-walled latewood fibres or marginal parenchyma band (Figure 1-1); wood is ring to semi-ring porous; vessels are solitary,infrequently in multiples and clusters; outline of vessels is round,oval or irregular,with 59(19-85)μm in tangential diameter for earlywood vessels; vessel element length is 152(86-220)μm.

Figure 1. Anatomical structure of Calligonum mongolicum. 1. TS. Growth ring boundary is distinct, marked by parenchyma bands; 2. TLS. Rays are 1-5-seriate; 3. RLS. Rays consist of procumbent cells, with square marginal cells; perforation plate is simple; 4. RLS. Intervessel pitting is alternate, vestured; 5. TLS. Vessel-parenchyma pitting is similar to intervessel pitting, alternate; 6. TLS. Helical thickening presents in narrow vessel element. Abbreviation: TS: transverse section; TLS: tangential longitudinal section; RLS: radial longitudinal section. LM: light microscope; SEM: scanning electronic microscope.
Ray is 388(180-1140)μm in height and ray frequency is 7(5-9)/mm; rays are uni- or 2-5-seriate (Figure 1-2) and distend at growth ring boundary; body ray cells are procumbent,with square marginal cells (Figure 1-3); pits are shown on the tangential wall and radial wall of ray cell,round,oval to elongate.
Perforation plate is exclusively simple (Figure 1-3). Intervessel pitting is alternate,some vestured; pit apertures are round to oval,elongated,slit-like or partly coalescent (Figure 1-4); intervessel pit diameter is 2.2(1.7-2.8)μm; vessel-parenchyma pitting is similar to the intervessel pitting in size and shape (Figure 1-5).
Distinct helical thickenings are throughout narrow vessels and parts of wide vessels (Figure 1-6). 568(334-812)μm long nonseptate fibres present with simple to bordered,slit-like pits.
Axial parenchyma abundantly present,paratrcheal to scanty paratracheal or marginal bands; fusiform parenchyma cells present and consists of 2-4 cells per parenchyma strand. This feature is different from Polygonum spp. without axial parenchyma [2].
3.2 Horizontal variation in vessel element length and fibre length
Vessel element length and fibre length within one sample are showed in Figure 2 . It is shown that there is a slightly increase of fibre length as the diameter from the pith increases. However,the vessel element length remains more or less constant from pith to bark. This pattern is very common in trees and is similar to the previous study on horizontal variations [5-8].

Figure 2. Variation in vessel element length and fibre length in Calligonum mongolicum.
4. Discussions
4.1 Ecological wood anatomy perspectives and significant functions
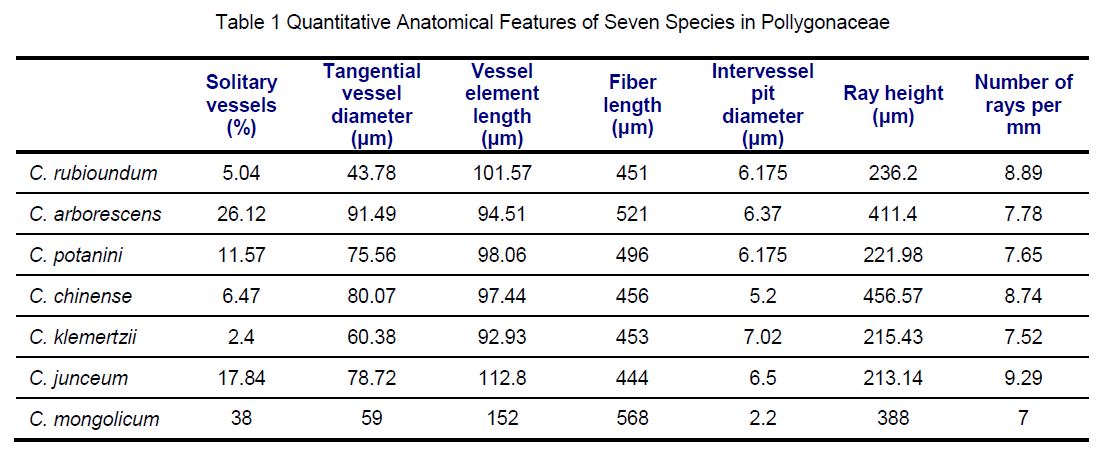
This species showed similar wood structure to that species published in the same genus[1],but there were few quantitative differences observed between them (Table1). Wood or secondary xylem provides a complex tissue for water transport,mechnical strength,and for metabolic processes such as storage and mobilization of reserve carbohydrates and lipids [9-12]. The species occuring in habitats subject to high water stress,their anatomical features of both xylem and leaf show adaptations correlated to environmental extremes. Leaf surface reduction with thick cuticles,photosynthesizing by green stems and cutinization of the outer walls in leaf epidermis enable plants to withstand dry climatic periods [13,14]. Ecological and evolutionary trends in vessel diameter,perforation plate type,vessel frequency,vessel member length,total vessel length,and fibre type have all been discussed in terms of their input to the safety and efficiency of water transport [9,15]. The tendencies are for vessel members to become shorter and narrower as the aridity increases to prevent collapse of vessels under high negative pressures [16] and vessels towards grouping in arid environments [9,17-21]. Vessels mainly solitary or few vessel grouping,narrow and numerous could lead to greater conductive safety because it renders the inactivation of any vessel less harmful by enabling the water transport to be transferred to an adjacent vessel [22].
Both efficiency or maximal conductivity and safety are strongly related to vessel diameter and vessel frequency. Increased vessel diameter increases efficiency of water conduction dramatically,but at the same time it decreases safety. However,ring-porosity and presence of different vessel size classes in general are of importance for the combined efficiency and safety of xylem sap transport at different times in or throughout the growing seasons [23]. The gradually decreased vessel diameter from earlywood to latewood allows for optimal transport efficiency by wide vessels and provides great conductive safety through the narrow latewood vessels [24-26].
Apart from these quantitative characters,the qualitative characters show ecological correlations.
Some narrow vessels in these species have coarse helical thickenings,which increase cell wall strength to withstand high pressures or enlarge wall surface to promote water bonding to the surface [27-28]. In this study,all species show helical thickenings,together with vessel groups and tracheids,which are associated with greater conductive safety in arid environments.
4.2 Horizontal variations in vessel element length and fibre length
Variations in cell length and volume have been discussed for long time due to their marked effect on product quality and the utilization of wood [29]. Particularly,fibre length is considered as one of the more important indicators of wood quality,which related to the mechanical strength and longitudinal shrinkage of wood. It is intensively studied within individual annual rings,from the base to the top of tree,among different rings within one tree,among species,and even on different sides of tree in relation to the sun and temperature [30].
Within-tree pattern of each species in each environment is different [29]. Commonly,previous studies on wood fibre length in the axial variation agreed with the tendency that fibre length increases more often to a point well up the bole and then decrease [7,31-35]. Other researchers have reported a constant length or a decrease in fibre length with height. Most patterns of fibre length in horizontal direction increasing with age in rings near the pith,following by a more gradual increase until a maximum reached have been reported for both hardwood and softwood [6-8],but there are other trends,such as,the cell length is constant,or considerable fluctuations or decrease with age. In present study,fibre length and vessel element length of selected species are shown unremarkable fluctuations or nearly constant. This trend is different from general conclusions that Furukawa et al. [36,37] has obtained from the study on 71 species hardwoods.
Vessel element length and fibre length is dictated by fusiform cambial initial cell length [10]. Additionally,fibre dimensions are determined by the dimensions of the cambial fusiform cells from which they are derived from and by process that occur during cell differentiation [33,38]. In species with non-storied cambium,the increase of fibre length is explained on the basis of the increase in the length of cambial initials with increasing cambial age [39]. In this study,most species have regular or irregular storied structure,and nearly constant length could result similarly from a retarded production from the cambial initials and lower degree of intrusive growth. The difference in range of fibre length is attribute to differences in age of wood and to the different species.
References
- Ma R-J,Wu S-M&Wang F-C,Studies on anatomy and desert adaptability of stem secondary xylem in 6 species of calligonum . Acta Botanica Sinica,36: 55-60 (1994 in Chinese).
- Schweingruber F. H.,Anatomy of European Woods,Haupt,259 (1990).
- Exley,R. R.,B. A. Meylan & B. G. Butterfield,A technique for obtaining cut surfaces on wood samples prepared for the scanning electron microscope,J. Microscopy,110:75-78 (1977).
- IAWA COMMITTEE,IAWA list of microscopic features for hardwood identification,IAWA Bull. n. s.,10: 219-332 (1989).
- Furukawa I.,S. Fukutani & J. Kishimoto,Characteristics of the variation of wood quality within Keyaki (Zelkova aerrata Makino) trees—horizontal variations of ring width,fiber length,vessel element length,specific graity and longitudinal compression strength. Hardwood Research. 5: 197-206 (1989).
- Stringer J. W. & J. R. Olson,Radial and vertical variation in stem properties of juvenile black locust (Robinia pseudoacacia). Wood and Fibre Length,19 (1): 59-67 (1987).
- Tayor F. W.,Variations in the size and proportions of wood elements in Yellow-Poplar trees. Wood Science and Technology,2:153-165 (1968).
- Tayor F. W.,Property variation within stems of selected hardwoods growing in Mid-south,Wood Science,11 (3): 193-199 (1979).
- Baas P.,Ecological patterns in xylem anatomy. In: T. J. Givnish (ed.),On the economy of plant form and function. Proceeding (11): 327-352(1986). Cambridge Univ. Press,Cambridge,London,New York,New Rochelle,Melbourne,Sydney.
- Carlquist S.,Comparative wood anatomy. Systematic,ecological,and evolutionary aspects of dicotyledon wood,Springer-Verlag,Berlin (1988).
- Zimmermann M. H.& C. L. Brown,Trees: Structure and function. Springer-Verlag,New York. (1977).
- Zimmermann M. H.,Xylem structure and ascent of sap. Springer-Verlag,Berlin,Heidelberg,New York,Tokyo. (1983).
- Lindorf H.,Wood and leaf anatomy in Sessea corymbiflora from an ecological perspective,IAWA J. 18 (2): 157-168 (1997).
- Villagra P. E. & F. A. Roig Junent,Wood structure of Prosopis alpataco and P. argentina growing under different edaphic conditions,IAWA J. 18 (1): 37-51 (1997).
- Zimmermann M. H. & C. L. Brown,Trees,structure and function. Springer-Verlag,Berlin,Heidelberg,New York. (1971).
- Barajas-Morales J.,Wood structural differences between trees of two tropical forests in Miexico. IAWA Bull. n. s. 6 (4): 355-364 (1985).
- Carlquist S.,Wood anatomy of Compositae: a summary with comments on factors controlling wood evolution. Aliso,6: 25-44 (1966).
- Carlquist S. & D. A. Hoekman,Ecological wood anatomy of the woody southern Californian flora,IAWA Bull. n. s.,6 (4): 319-347 (1985).
- Fahn. A.,E. Werker & P. Baas,Wood anatomy and identification of trees and shrubs from Israel and adjacent regions. Israel Acadamy of Sciences,Jerusalem,Israel (1986).
- Lindorf H.,Eco-anatomical wood features of species from a very tropical forest. IAWA J. 15 (4): 361-376 (1994).
- Zhong Y.,P. Baas & E. A. Wheeler,Wood anatomy of trees and shrubs from China. . Ulmaceae. IAWA ÃÆâÃâââ¬Â¦ÃâãBull. n. s. 13 (4): 419-453 (1992).
- Carlquist S.,Vessel grouping in dicotyledon wood: significance and relationship to imperforate tracheary elements,Aliso,10: 505-525 (1984).
- Baas P. & F. H. Schweingruber,Ecological trends in the wood anatomy of trees,shrubs and climbers from Europe. IAWA Bull. n. s. 8 (3): 245-274 (1987).
- Baas P.,M. Esser.,M. E. T. van den Western & M. Zandee,Wood anatomy of Oleaceae,IAWA Bull. n. s. 9: 103-182 (1988).
- Woodcock D. W.,Occurrence of woods with a gradation in vessel diameter across a ring. IAWA J. 15 (4): 377-385 (1994).
- Zimmermann M. H.,Functional xylem anatomy of angiosperm trees. In: new perspectives in wood anatomy,59-70. Nijhoff/Junk,The Hague,Boston (1982).
- Carlquist S.,Ecological strategies of xylem evolution,Univ. Calif. Press,Berkeley,Los Angeles,London (1975).
- Carlquist S.,Wood anatomy of Illicium (Illiciaceae): phylogenetic,ecological,and functional interpretation,Amer. J. Bot.,69: 1587-1598 (1982).
- Zobel B. J. & J. P. van Buijtenen,Wood variation,its causes and control. Springer-Verlag. 1-32 (1989).
- Slunder E. R.,Variation in wood specific gravity of yellow poplar in the southern Appalachians. Wood S. 5: 132-138 (1972).
- Bhat K. M.,K. V. Bhat & T. K. Dhamodaran,Wood density and fibre length of Eucalyptus grandis grown in Kerala,India. Wood and Fibre Sci. 22: 54-61 (1990).
- Olson J. R. & S. B. Carpenter,Specific gravity,fibre length,and extractive content of young Paulownia. Wood and Fibre Science,17 (4): 428-438 (1985).
- Ridoutt B. G. & R. Sands,Within-tree variation in cambial anatomy and xylem cell differenciation in Eucalyptus globules. Trees,8: 18-22 (1993).
- Wilkes J.,Variations in wood anatomy within species of Eucalypus. IAWA Bull. n. s. 9: 13-23 (1988).
- Yamada M.,T. Sakuno,I. Furukawa & J. Kishimoto,Wood quality of small hardwoods() within tree ÃÆâÃâââ¬Â¦Ãâãvariation of fibre length in Niseakashia (Robinia pseudoacacia L). Hardwood Reasearch,3:
- Furukawa I.,T. Nakayama,T. Sakuno & J. Kishimoto,Wood quality of small hardwoods,2. Horizontal variations in the length of fibres and vessel elements in seventy-one species of small hardwoods. Hardwood Research. 2: 104-134 (1983a).
- Furukawa I.,T. Nakayama,T. Sakuno & J. Kishimoto,Wood quality of small hardwoods,3. Horizontal variations in the length of fibres and vessel elements in trees with storeyed and non-storeyed wood. Bull. Fac. Agric. Tottori Univ. 35: 42-49 (1983b).
- Ridoutt B. G. & R.Sands,Quantification of the process of secondary xylem fibre development in Eucalyptus globules at two height levels. IAWA J. 15: 417-424 (1994).
- Heinowicz A. & Z. Heinowicz,Variations of length of vessel members and fibres in the trunk of Robinia pseudoacacia. Acta Societatis Botanicorrum Poloniae,28 (3): 453-460 (1959).


Open Access Journals
- Aquaculture & Veterinary Science
- Chemistry & Chemical Sciences
- Clinical Sciences
- Engineering
- General Science
- Genetics & Molecular Biology
- Health Care & Nursing
- Immunology & Microbiology
- Materials Science
- Mathematics & Physics
- Medical Sciences
- Neurology & Psychiatry
- Oncology & Cancer Science
- Pharmaceutical Sciences